{kind=link}

The capacity to respond to the presence of lactose was always there. The genes for the three induced enzymes are part of the genome of the cell. But until lactose was added to the culture medium, these genes were not expressed.
The most direct way to control the expression of a gene is to regulate its rate of transcription; that is, the rate at which RNA polymerase transcribes the gene into molecules of messenger RNA (mRNA).
| Link to a general discussion of gene transcription. |
Gene transcription begins at a particular nucleotide shown in the figure as "+1". RNA polymerase actually binds to a site "upstream" (i.e., on the 5' side) of this site and opens the double helix so that transcription of one strand can begin.
 The binding site for RNA polymerase is called the promoter. In bacteria, two features of the promoter appear to be important:
The binding site for RNA polymerase is called the promoter. In bacteria, two features of the promoter appear to be important:
The exact DNA sequence between the two regions does not seem to be important.
Each of the three enzymes synthesized in response to lactose is encoded by a separate gene. The three structural genes (so named because they encode a product) are arranged in tandem on the bacterial chromosome.
| The lac operon. In the absence of lactose, the repressor protein encoded by the I gene binds to the lac operator and prevents transcription. Binding of lactose (and its relatives) to the repressor causes it to leave the operator. This enables RNA polymerase to transcribe the three structural genes of the operon. The single mRNA molecule that results is then translated into the three proteins. |
|
The lac repressor binds to a specific sequence of two dozen nucleotides called the operator. Most of the operator is downstream of the promoter. When the repressor is bound to the operator, RNA polymerase is unable to proceed downstream with its task of gene transcription.
The operon is the combination of the
The gene encoding the lac repressor is called the I gene. It happens to be located just upstream of the lac promoter. However, its precise location is probably not important because it achieves its effect by means of its protein product, which is free to diffuse throughout the cell. And, in fact, the genes for some repressors are not located close to the operators they control.
The lac repressor is made up of four identical polypeptides (thus a "homotetramer"). Part of the molecule has a site (or sites) that enable it to recognize and bind to the 24 base pairs of the lac operator. Another part of the repressor contains sites that bind to lactose. When lactose unites with the repressor, it causes a change in the shape of the molecule, so that it can no longer remain attached to the DNA sequence of the operator. Thus, when lactose is added to the culture medium,Hardly does transcription begin before ribosomes attach to the growing mRNA molecule and move down it to translate the message into the three proteins. You can see why punctuation codons - UAA, UAG, or UGA - are needed to terminate translation between the portions of the mRNA coding for each of the three enzymes.
This mechanism is characteristic of prokaryotes, but differs in three respects from that found in eukaryotes:
|
As mentioned above, the synthesis of tryptophan from precursors available in the cell requires 5 enzymes. The genes encoding these are clustered together in a single operon with its own promoter and operator. In this case, however, the presence of tryptophan in the cell shuts down the operon. When Trp is present, it binds to a site on the Trp repressor and enables the Trp repressor to bind to the operator. When Trp is not present, the repressor leaves its operator, and transcription of the 5 structural genes begins.
| Stereo view of the tryptophan repressor (right side of each panel) bound to its operator DNA (left side). The repressor is a dimer of two identical polypeptides (on either side of the horizontal red line). Binding to DNA occurs only when a molecule of tryptophan (red rings) is bound to each monomer of the repressor. You may be able to fuse the images be holding a stiff paper or cardboard between the views so that your left eye sees only the left-hand image, your right eye the right-hand image. Persevere; the results are worth it!. (Image courtesy of P. B. Sigler.) |  |
The usefulness to the cell of this control mechanism is clear. The presence in the cell of an essential metabolite, in this case tryptophan, turns off its own manufacture and thus stops unneeded protein synthesis.
As its name suggests, repressors are negative control mechanisms, shutting down operonsHowever, CAP can bind to DNA only when cAMP is bound to CAP. so when cAMP levels in the cell are low, CAP fails to bind DNA and thus RNA polymerase cannot begin its work, even in the absence of the repressor.
So the lac operon is under both negative (the repressor) and positive (CAP) control.
Why?
It turns out that it is not simply a matter of belt and suspenders. This dual system enables the cell to make choices. What, for example, should the cell do when fed both glucose and lactose? Presented with such a choice, E. coli (for reasons about which we can only speculate) chooses glucose. It makes its choice by using the interplay between these two control devices.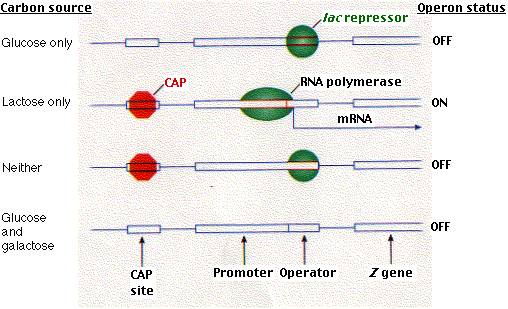
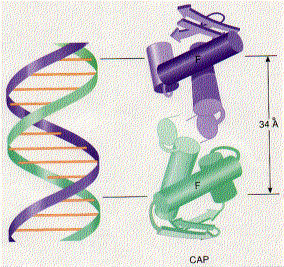
CAP consists of two identical polypeptides (hence it is a homodimer). Toward the C-terminal, each has two regions of alpha helix with a sharp bend between them. The longer of these is called the recognition helix because it is responsible for recognizing and binding to a particular sequence of bases in DNA.
The graphic shows a model of CAP. The two monomers are identical. Each monomer recognizes a sequence of nucleotides in DNA by means of the region of alpha helix labeled F. Note that the two recognition helices are spaced 34Å apart, which is the distance that it takes the DNA molecule (on the left) to make precisely one complete turn.

The recognition helices of each polypeptide of CAP are, of course, identical. But their orientation in the dimer is such that the sequence of bases they recognize must run in the opposite direction for each recognition helix to bind properly. This arrangement of two identical sequences of base pairs running in opposite directions is called an inverted repeat.
| Welcome&Next Search |