Antigen Presentation
Antigens are macromolecules that elicit an immune response in the body. The most common antigens are proteins and polysaccharides.
It will be helpful to distinguish between
- antigens that enter the body from the environment; these would include
- inhaled macromolecules (e.g., proteins on cat hairs that can trigger an attack of asthma in susceptible people)
- ingested macromolecules (e.g., shellfish proteins that trigger an allergic response in susceptible people)
- molecules that are introduced beneath the skin (e.g., on a splinter or in an injected vaccine)
- antigens that are generated within the cells of the body; these would include
- proteins encoded by the genes of viruses that have infected a cell
- aberrant proteins that are encoded by mutant genes; such as mutated genes in cancer cells
In all cases, however, the initial immune response to any antigen absolutely requires that the antigen be recognized by a T lymphocyte ("T cell"). The truth of this rule is clearly demonstrated in AIDS: the infections (viral or fungal or bacterial) that so often claim the life of AIDS patients do so when the patient has lost virtually all of his or her CD4+ T cells.
The two categories of antigens are processed and presented to T cells by quite different mechanisms.
First Group: Exogenous antigens
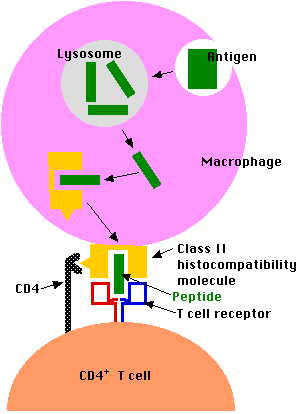
Exogenous antigens (inhaled, ingested, or injected) are taken up by "professional" antigen-presenting cells; that is, cells for which this function is their primary purpose. These include:
- phagocytic cells like macrophages and dendritic cells
- B lymphocytes ("B cells"); which are responsible for producing antibodies against the antigen.
Professional antigen-presenting cells
- engulf the antigen by endocytosis
- degrade it into fragments (e.g. break an antigenic protein into short peptides)
- display these peptides at the surface of the cell nestled within a
- class II histocompatibility molecule
- here they may be recognized by CD4+ T cells
Second Group: Endogenous antigens
Antigens that are generated within a cell (e.g., viral proteins in any infected cell) are
- degraded into fragments (e.g., peptides) within the cell and
- displayed at the surface of the cell nestled within a
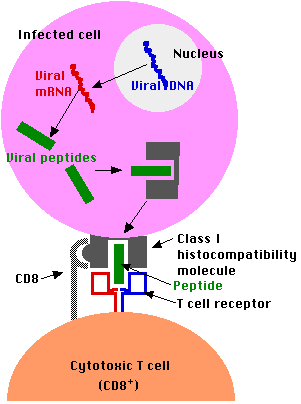
- class I histocompatibility molecule.
- Here they may be recognized by CD8+ T cells.
- Most CD8+ T cells are cytotoxic.
- They have the machinery to destroy the infected cell (often before it is able to release a fresh crop of viruses to spread the infection).
Now for more details.
The Class I Pathway
Class I histocompatibility molecules are transmembrane proteins expressed at the cell surface. Like all transmembrane proteins, they are synthesized by ribosomes on the rough endoplasmic reticulum (RER) and assembled within the lumen of the RER.
There are three subunits in each class I histocompatibility molecule:
- the transmembrane polypeptide (called the "heavy chain")
- the antigenic peptide
- beta-2 microglobulin
All of these must be present within the lumen of the RER if they are to assemble correctly and move through the Golgi apparatus to the cell surface.
The Problem: proteins encoded by the genes of an infecting virus are synthesized in the cytosol. How to get them to the RER?
The Solution: TAP (= transporter associated with antigen processing).
- Viral proteins in the cytosol are degraded (probably by proteasomes) into viral peptides.
- The peptides are picked up by TAP proteins and transported from the cytosol into the RER where they assemble with
- the transmembrane polypeptide and beta-2 microglobulin.
- This trimolecular complex then moves through the Golgi apparatus and is inserted in the plasma membrane.
- The complex can be bound by a T cell with

- a receptor (TCR) able to bind the peptide and flanking portions of the histocompatibility molecule (the hot dog in the bun) and
- CD8 molecules that bind the CD8 receptor (shown above as a gray hemisphere) on the histocompatibility molecule.
Class II histocompatibility molecules consist of
- two transmembrane polypeptides and
- a third molecule nestled in the groove they form.
All three components of this complex must be present in the RER for proper assembly.
But antigenic peptides are not transported to the RER, so a protein called the invariant chain ("Ii") temporarily occupies the groove.
The steps:
- The two chains of the class II molecule are inserted into the membrane of the RER.
- They bind (in their groove) one molecule of invariant chain.
- This trimolecular complex is transported through the Golgi apparatus and into vesicles called lysosomes.
Meanwhile,
- Foreign antigenic material is engulfed by endocytosis forming endosomes.
- These also fuse with lysosomes.
Then,
- The antigen is digested into fragments.
- The invariant (Ii) chain is digested.
- This frees the groove for occupancy by the antigenic fragment.
- The vesicles move to the plasma membrane and the complex is displayed at the cell surface.
- The complex can be bound by a T cell with
- a receptor (TCR) able to bind the peptide and flanking portions of the histocompatibility molecule (the hot dog in the bun) and
- CD4 molecules that bind the CD4 receptor (shown above as a yellow triangle) found on all class II histocompatibility molecules.
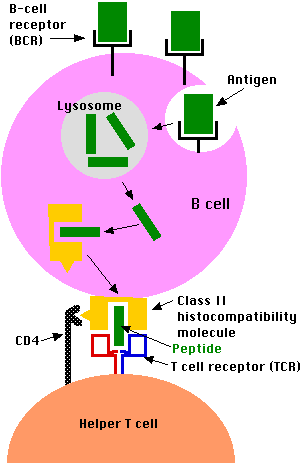
B lymphocytes process antigen by the class II pathway.
However, antigen processing by B cells differs from that of phagocytic cells like macrophages in crucial ways:
- B cells engulf antigen by receptor-mediated endocytosis
- The B cell receptors for antigen (BCRs) are antibodies anchored in the plasma membrane.
- The affinity of these for an epitope on an antigen may be so high that the B cell can bind and internalize the antigen when it is present in body fluids in concentrations thousands of times smaller than a macrophage would need.
- The remaining steps of antigen processing occur by the same class II pathway described above for macrophages producing
- fragments of antigen displayed at the cell surface nestled in the groove of class II histocompatibility molecules.
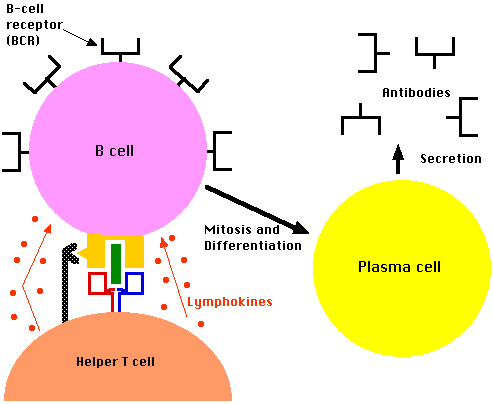
- A CD4+ T cell that recognizes the displayed antigen is stimulated to release lymphokines.
- These, in turn, stimulate the B cell to enter the cell cycle.
- Because of the part they play in stimulating B cells, these CD4+ T cells are called Helper T cells ("Th").
- The B cell grows into a clone of cells (called plasma cells)
- These synthesize receptors (BCRs) with the identical binding site for the epitope but without the transmembrane tail.
- These antibodies are secreted into the surroundings.
23 May 1999
{kind=link}