The human genome is presently estimated to contain from 70 thousand to as many as 100 thousand (105) genes (in a few years we should know the number exactly). The number of T-cell receptors for antigen (TCRs) that we make is estimated at 2.5 x 107; the number of different kinds of antibody molecules (BCRs) is probably about the same.
| Link to discussion of the structure of BCRs and TCRs. |
How could 105 genes encode 2.5 x 107 different TCRs and the same number of different BCRs?
The answer: each receptor chain (heavy, kappa, and lambda chains for BCRs; alpha and beta chains for TCRs) is encoded by several different gene segments.
The genome contains a pool of gene segments for each type of chain. Random assortment of these segments makes the largest contribution to receptor diversity.
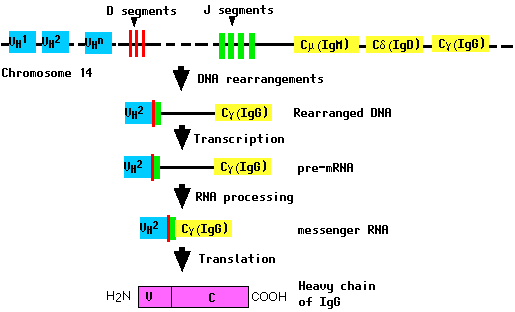
During the differentiation of the B cell (and long before any possible encounter with an antigen), the DNA in this locus is cut and recombined to make an intact gene for the heavy chain. This gene can then be transcribed into mRNA, which is, in turn, translated into the heavy (H) polypeptide chain.
The random assembly of 50 VH, 29 DH, and 6 JH gene segments provides a minimum of 8.7 x 103 different possible combinations.
But the possibilities of antibody V region diversity turn out to be greater than that. The recombination process is not precise.| Antibodies (BCRs) | Gene Segments | Combinations | |
|---|---|---|---|
| V-kappa | 32 | ||
| J-kappa | 5 | ||
| N regions | 100 | 1.6 x 104 kappa chains | |
| VH | 50 | ||
| DH | 29 | ||
| JH | 6 | ||
| N regions | 100 | 8.7 x 105 H chains | |
| Any H chain with any kappa L chain | 14 x 109 | ||
The diversifying mechanisms described above take place before the B cell encounters antigen. After a B cell encounters antigen, it may begin mitosis, growing into a clone of cells synthesizing the same BCR (and, eventually, secreting antibodies with the same binding site). As in any dividing cell, point mutations occur. While most are deleterious, those occurring in the DNA encoding the hypervariable regions MAY generate a binding site with increased affinity for its epitope. These are favorable mutations, and the "sub-clone" in which they occur tends to be favored and may replace the ancestral clone. The result is affinity maturation, the production of antibodies of ever-increasing affinity for the antigen.
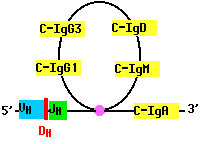
As B cells grow into a clone in response to antigen, they may rearrange their DNA once again. For example, a B cell that has assembled a complete gene for the H chain of IgM (mu), may cut the gene on the 3´ side of the assembled V-region segments and move the assembly to the 5´ side of another of its C-region gene segments. Now the cell begins to make a different kind of antibody, such as IgG or IgA. But the antigen-specificity of the antibody remains the same because the N-terminal of the H chain remains unchanged (as does the L chain).
This switching mechanism enables the body to produce antibodies with different effector functions; that is, different means of dealing with the same antigen.
The ability of a B cell to switch CH gene segments depends on its receiving help from helper T cells.
The most abundant T cells in the blood express a receptor for antigen (TCR) that is a heterodimer of two chains designated alpha and beta. Each of these is encoded by a gene assembled from V, D, J, and C gene segments. Like BCRs, there are multiple variants of these gene segments arranged in clusters:
| T cell receptors (TCRs) | Gene segments | Combinations | |
|---|---|---|---|
| V-alpha | 50 | ||
| J-alpha | 50 | ||
| N regions | 100 | 2.5 x 105 alpha chains | |
| V-beta | 20 | ||
| J-beta | 13 | ||
| D-beta | 2 | ||
| N regions | 100 | 5.2 x 104 beta chains | |
| Any alpha with any beta chain | 1.3 x 1010 | ||
| Welcome&Next Search |