Embryonic Development: Getting Started
In animals, one can usually distinguish 4 stages of embryonic development.
- Cleavage
- Patterning
- Differentiation
- Growth
Cleavage
Mitosis and cytokinesis of the zygote, an unusually large cell, produces an increasing number of smaller cells, each with an exact copy of the genome present in the zygote. However, the genes of the zygote are not expressed at first. The activities of cleavage are controlled by the mother's genome; that is, by mRNAs and proteins she deposited in the unfertilized egg. Cleavage ends with the formation of a blastula.
Patterning
During this phase, the cells produced by cleavage organize themselves in layers and masses, a process called gastrulation. The pattern of the future animal appears: - front and rear (the anterior-posterior axis)
- back side and belly side (its dorsal-ventral axis)
- left and right sides.
There is little visible differentiation of the cells in the various layers, but probes for cell-specific proteins reveal that different groups of cells have already started on specific paths of future development.
Gastrulation forms three major "germ layers": ectoderm, mesoderm, and endoderm. By gastrulation, the genes of the zygote genome are being expressed.
Differentiation
In time, the cells of the embryo differentiate to form the specialized structures and functions that they will have in the adult. They form neurons, blood cells, skin cells, muscle cells, etc., etc. These are organized into tissues, the tissues into organs, the organs into systems.
Growth
After all the systems are formed, most animals go through a period of growth. Growth occurs by the formation of new cells and more extracellular matrix.
The Problem
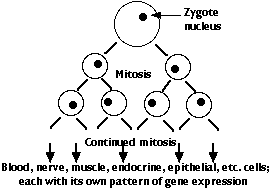 The genome of the zygote contains all the genes needed to make the hundreds of different types of cells that will make up the complete animal. There are two major categories of these genes:
The genome of the zygote contains all the genes needed to make the hundreds of different types of cells that will make up the complete animal. There are two major categories of these genes:
- "housekeeping" genes = those that encode the RNAs and proteins needed by all kinds of cells. Examples:
- genes for tRNAs, rRNAs
- enzymes of glycolysis.
- tissue-specific genes = those that encode mRNAs and hence proteins that are used by one or a few specific kinds of cell. Examples:
- genes for hemoglobin expressed in the precursors of red blood cells
- the gene for insulin expressed in the beta cells of the islets of Langerhans
However, every cell descended from the zygote has been produced by mitosis and thus contains the complete genome of the organism (with a very few exceptions).
Two pieces of evidence:
- Dolly. Dolly is the sheep that was formed by inserting a nucleus from a single cell of an adult sheep into an enucleated sheep egg. She proves that the cell from the adult had lost none of the genes needed to build all the tissues of a sheep.
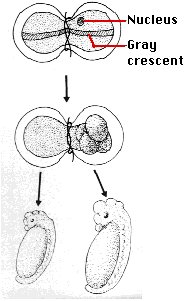
- Spemann's egg-tying experiments. Many years earlier, the German embryologist Hans Spemann demonstrated the same truth. He used strands of baby hair to tie loops around fertilized salamander eggs. Although the egg half with the nucleus began cleaving normally, the other side did not begin cleavage until a nucleus finally slipped through the knot. So long as the egg was tied so that both halves contained some of the gray crescent, the second half began normal cleavage and ultimately produced a second tadpole. Even after 5 mitotic division of the zygote nucleus (the 32-cell stage), the entire genome was still available in each descendant nucleus.
SO,
How do the various differentiated cells come to express the appropriate subset of tissue-specific genes and not all the genes of the genome?
The Solution
- A fertilized egg is much larger than the normal cells of an animal's body. Some (e.g., a hen's egg) are truly huge. The frog egg has a volume 1.6 millions times larger than a typical frog cell. The eggs of mammals are smaller, but even they are larger than their descendant cells will be. The photo (courtesy of L. M. Beidler) is of a 16-cell frog embryo. This mass of cells is no larger than the original egg.

- The cytoplasm of the fertilized egg is NOT HOMOGENEOUS. It contains gradients of mRNAs and proteins. These are the products of the mother's genes and were deposited in the egg by her.
- Cleavage of the fertilized egg partitions it into thousands of cells of normal size. Each contains a nucleus descended from the zygote nucleus.
- The genes that are expressed by the nucleus in a given cell are regulated by the molecules, mostly transcription factors, found in the cytoplasm surrounding that nucleus.
- Once a cell-specific pattern of gene expression is launched, that cell may release molecules that regulate the genes of nearby cells.
- In this way, the foundation is laid for the building of an organism with hundreds of types of differentiated cells each in its correct location and performing its correct functions.
First Example: Xenopus
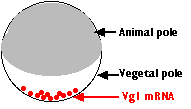
- During egg formation, molecules of mRNA encoding the protein Vg1 collect at the vegetal pole of the cell.
- Cells that form there during cleavage translate the mRNA into the Vg1 protein.
- This is secreted by the vegetal pole cells and induces the cells above them to start down the path to becoming mesoderm.
- Some of those cells will, in turn, become the Spemann organizer.
- Later, the Spemann organizer will secrete molecules that induce the ectodermal cells above them to develop into the tissues of the brain and spinal cord.


The photo on the left (courtesy of Douglas Melton, in whose lab the work was done) shows the tadpole that developed from a frog embryo that had been injected with mRNA for activin (a growth factor similar to Vg1 ). At the 32-cell stage of cleavage, a single cell - that would normally have gone on to form part of the belly of the animal - was injected with activin mRNA. A two-headed tadpole resulted (arrow). (A normal tadpole is shown on the right for comparison.)
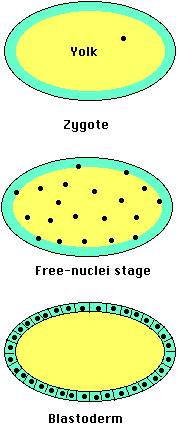
In Drosophila and other insects, cleavage involves repeated mitosis but without cytokinesis. So the daughter nuclei remain suspended within the single egg compartment. After several thousand nuclei have been formed, they migrate to the margins of the egg. Only then do plasma membranes form around each nucleus forming true cells . But, as for Xenopus, the genes that will be expressed by those cells are regulated by the cytoplasmic constituents they found themselves surrounded by. And that, again like Xenopus, is determined by where the mother deposited those molecules when she made the egg.
Bicoid
For example, Drosophila eggs have a gradient of mRNA transcribed from a gene designated bicoid (bcd). The transcripts are transported into the egg by "nurse" cells surrounding it. The result is a concentration gradient of bicoid mRNA extending from a high level at the anterior of the egg to a low level at the posterior. The signal for the nurse cells to deposit bicoid mRNA at the anterior is located in the 3´ region of the bicoid gene.
After fertilization, the mRNAs are translated into bicoid protein. High levels of the protein lead to the formation of the head of the larva.
Conversely, the posterior of the egg has a high concentration of mRNA encoding the nanos protein, which is needed to form the structures of the tail of the larva.
Three demonstrations:
1.
Remove some of the bicoid-rich cytoplasm from the anterior of the fertilized egg and replace it with nanos-rich cytoplasm from another egg.
The result: a larva (non-viable) with a tail at each end.
2.
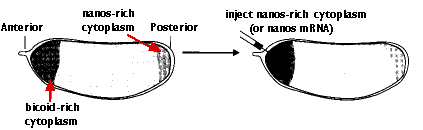
Inject the anterior of the fertilized egg with nanos mRNA. The result: another double-posterior larva.
3.
Make female fruit flies that are transgenic for a recombinant gene containing: - the structural gene for nanos
- coupled to the 3´ anterior-directing signal of the bicoid gene.
This causes nanos mRNA, instead of bicoid mRNA, to be deposited in the anterior of her eggs. The result: more double-posterior larvae (on the left).


A normal larva is shown on the right. The bright object at the right end of the normal larva and at both ends of the double posterior larva is the tip of the tail. These micrographs are courtesy of Elizabeth Gavis and Ruth Lehmann, in whose lab the third demonstration was performed.
What comes next?
Development in Xenopus and Drosophila (and probably that of animals in general) passes through three rather different (although often overlapping) phases:
- establishing the main axes (dorsal-ventral; anterior-posterior; left-right). This is done by gradients of mRNAs and proteins encoded by the mother's genes and placed in the egg by her. It has been discussed here.
- establishing the main body parts such as the notochord and central nervous system in vertebrates
and the segments in Drosophila
These are run by genes of the zygote itself.
- filling in the details; that is, building the various organs of the animal. (Our example will include the wings, legs, and eyes of Drosophila.)
27 May 1999