
The inheritance of one pair of factors (genes) is independent of the inheritance of the other pair.
Today we know that this rule holds only if two conditions are met:Mendel was lucky in that every pair of genes he studied met one requirement or the other. The table shows the chromosome assignments of the seven pairs of alleles that Mendel studied. Although all of these genes showed independent assortment, several were, in fact, syntenic with three loci occurring on chromosome 4 and two on chromosome 1. However, the distance separating the syntenic loci was sufficiently great that the genes were inherited as though they were on separate chromosomes.
| Trait | Phenotype | Alleles | Chromosome |
|---|---|---|---|
| Seed form | round-wrinkled | R-r | 7 |
| Seed color | yellow-green | I-i | 1 |
| Pod color | green-yellow | Gp-gp | 5 |
| Pod texture | smooth-wrinkled | V-v | 4 |
| Flower color | purple-white | A-a | 1 |
| Flower location | axial-terminal | Fa-fa | 4 |
| Plant height | tall-dwarf | Le-le | 4 |
With the rebirth of genetics in the 20th century, it quickly became apparent that Mendel's second rule does not apply to many matings of dihybrids. In many cases, two alleles inherited from one parent show a strong tendency to stay together as do those from the other parent. This phenomenon is called linkage.
When the pollen of the first strain is dusted on the silks of the second (or vice versa), the kernels produced (F1) are all yellow and smooth. So the alleles for yellow color (C) and smoothness (Sh) are dominant over those for colorlessness (c) and shrunken endosperm (sh).
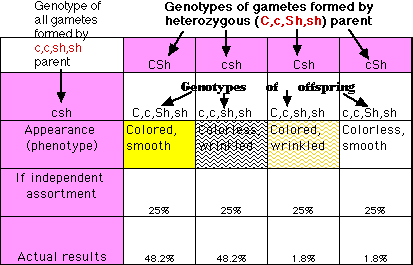 To simplify the analysis, mate the dihybrid with a homozygous recessive strain (ccshsh). Such a mating is called a test cross because it exposes the genotype of all the gametes of the strain being evaluated.
According to Mendel's second rule, the genes determining color of the endosperm should be inherited independently of the genes determining texture. The F1 should thus produce gametes in approximately equal numbers.
To simplify the analysis, mate the dihybrid with a homozygous recessive strain (ccshsh). Such a mating is called a test cross because it exposes the genotype of all the gametes of the strain being evaluated.
According to Mendel's second rule, the genes determining color of the endosperm should be inherited independently of the genes determining texture. The F1 should thus produce gametes in approximately equal numbers.
If the inheritance of these genes observes Mendel's second rule; i.e., shows independent assortment, union of these gametes should produce approximately equal numbers of the four phenotypes. But as the chart shows, there is instead a strong tendency for the parental alleles to stay together. It occurs because the two loci are relatively close together on the same chromosome. Only 3.6% of the gametes contain a recombinant chromosome.
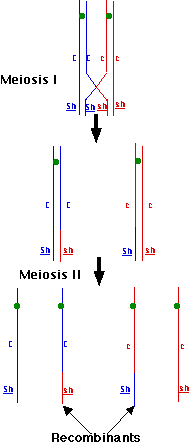
During prophase I of meiosis, pairs of duplicated homologous chromosomes unite in synapsis and then nonsister chromatids exchange segments during crossing over. It is crossing over that produces the recombinant gametes. In this case, whenever a crossover occurs between the locus for kernel color and that for kernel texture, the original combination of alleles (CSh and csh) is broken up and a chromosome containing Csh and one containing cSh will be produced.
The percentage of recombinants formed by F1 individuals can range from a fraction of 1% up to the 50% always seen with gene loci on separate chromosomes (independent assortment). The higher the percentage of recombinants for a pair of traits, the greater the distance separating the two loci. In fact, the percent of recombinants is arbitrarily chosen as the distance in centimorgans (cM), named for the pioneering geneticist Thomas Hunt Morgan. In our case, the c and sh loci are said to be 3.6 cM apart.
The procedure can be continued with another locus on the same chromosome.Test crossing a corn plant that is dihybrid for the C,c alleles and the alleles for bronze color (Bz, bz) produces 5% recombinants. So these two loci are 5 cM apart. However, is the bz locus on the same side of c as sh or is it on the other side?
The answer can be found by test crossing the dihybrid Shsh, Bzbz. If the percentage of recombinants is less than 5%, then bz must be on the same side of locus c as locus sh. If greater than 5%, it must be on the other side. In fact, the recombination frequency is less than 2%, telling us that the actual order of loci is c - sh - bz.
Mapping by linkage analysis is best done with loci that are relatively close together; that is, within a few centimorgans of each other. Why? Because as the distance between two loci increases, the probability of a second crossover occurring between them also increases. But a second crossover would undo the effect of the first and restore the parental combination of alleles. These would show up as nonrecombinants. Thus as the distance between two loci increases, the percentage of recombinants that forms understates the actual distance in centimorgans that separates them.
| View an example of multiple crossovers. |

There are other problems with preparing genetic maps of chromosomes.
Chromosome maps prepared by counting phenotypes are called genetic maps. They have been prepared for many eukaryotes, including corn, Drosophila, the mouse, and the tomato. A genetic map of chromosome 9 (the one that carries the C, Sh, and bz loci) of the corn plant (Zea mays) is shown on the right.
If one maps in small intervals from one end of a chromosome to the other, the total number of centimorgans often exceeds 100 (as you can see for chromosome 9). However, even for widely-separated loci, the maximum frequency of recombinants that can form is 50%. And this is also the frequency of recombinants that we see for genes independently assorting on separate chromosomes. So we cannot tell by simply counting recombinants whether a pair of gene loci is located far apart on the same chromosome or are on different chromosomes. As we saw above, several of Mendel's independently assorting traits are controlled by genes on the same chromosome but located so far apart that they are inherited as if they were located on different chromosomes.
Genes that are present on the same chromosome are called syntenic.
Obviously one cannot perform controlled matings of humans to map our chromosomes. However, it is possible to estimate map positions by examining linkage in several generations of relatives.
For example, blood samples from several large Mormon families in Utah, where all the members of at least three generations were alive to be sampled, have been collected and stored. These have already been used to establish genetic linkage relationships and will be available in the years ahead to study other human genes as they are identified.
| Using restriction fragment polymorphisms (RFLPs) to map the location of disease-causing genes in humans. |
Chromosome mapping by counting the number of recombinants produces a genetic map of the chromosome. But all the genes on the chromosome are incorporated in a single molecule of DNA. Genes are simply portions of the molecule (open reading frames or ORFs) encoding products that create the observed trait (phenotype). The rapid progress in DNA sequencing has produced complete genomes for many prokaryotes and two eukaryotes: budding yeast, Saccharomyces cerevisiae, and the roundworm, C. elegans.
| Table showing the organisms for which the complete genome is known. |
Having the complete sequence makes it possible to determine directly the order and spacing of the genes. Maps drawn in this way are called physical maps.
What is the relationship between the genetic map and the physical map of a chromosome? As a very rough rule of thumb, we shall assume that 1 cM on a chromosome encompasses 1 megabase (1 Mb = 106 bp) of DNA. But for the reasons mentioned above, this relationship is only approximate. Although the genetic maps of human females average 90% longer than the same maps in males, their chromosomes contain the same number of base pairs. So their physical maps are identical.
| Welcome&Next Search |