Apoptosis
For every cell, there is a time to live and a time to die.
There are two ways in which cells die:
- they are killed by injurious agents
- they are induced to commit suicide
Death by injury
Cells that are damaged by injury, such as by
- mechanical damage
- exposure to toxic chemicals
undergo a characteristic series of changes:
- they (and their organelles like mitochondria) swell (because the ability of the plasma membrane to control the passage of ions and water is disrupted)
- the cell contents leak out, leading to
- inflammation of surrounding tissues
Death by suicide
Cells that are induced to commit suicide:
- shrink
- have their mitochondria break down with the release of cytochrome c
- develop bubble-like blebs on their surface
- have the chromatin (DNA and protein) in their nucleus degraded
- break into small, membrane-wrapped, fragments which are usually
- engulfed by nearby phagocytic cells like macrophages
- do not induce inflammation
The pattern of events in death by suicide is so orderly that the process is often called programmed cell death or PCD. The cellular machinery of programmed cell death turns out to be as intrinsic to the cell as, say, mitosis.
Programmed cell death is also called apoptosis. (There is no consensus yet on how to pronounce it; some say APE oh TOE sis; some say uh POP tuh sis.)
Why should a cell commit suicide?
There are two different reasons.
1. Programmed cell death is as needed for proper development as mitosis is.
Examples:
- The resorption of the tadpole tail at the time of its metamorphosis into a frog occurs by apoptosis.
- The formation of the fingers and toes of the fetus requires the removal, by apoptosis, of the tissue between them.
- The sloughing off of the inner lining of the uterus (the endometrium) at the start of menstruation occurs by apoptosis.
- The formation of the proper connections (synapses) between neurons in the brain requires that surplus cells be eliminated by apoptosis
2. Programmed cell death is needed to destroy cells that represent a threat to the integrity of the organism.
Examples:
- Cells infected with viruses
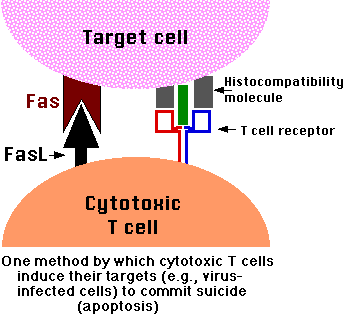
- One of the methods by which cytotoxic T lymphocytes (CTLs) kill virus-infected cells is by inducing apoptosis [diagram of the mechanism]. (And some viruses mount countermeasures to thwart it.)
- Cells of the immune system
- As cell-mediated immune responses wane, the effector cells must be removed to prevent them from attacking body constituents. CTLs induce apoptosis in each other and even in themselves. Defects in the apoptotic machinery is associated with autoimmune diseases such as lupus erythematosus and rheumatoid arthritis.
- Cells with DNA damage
- Damage to its genome can cause a cell
- to disrupt proper embryonic development leading to birth defects
- to become cancerous.
Cells respond to DNA damage by increasing their production of p53. p53 is a potent inducer of apoptosis. Is it any wonder that mutations in the p53 gene, producing a defective protein, are so often found in cancer cells (that represent a lethal threat to the organism if permitted to live)?
- Cancer cells
- Many agents used in cancer chemotherapy achieve their effect by inducing apoptosis in the cancer cells.
What makes a cell decide to commit suicide?
The balance between:
- the withdrawal of positive signals; that is, signals needed for continued survival
- the receipt of negative signals
Withdrawal of positive signals
The continued survival of most cells requires that they receive continuous stimulation from other cells and, for many, continued adhesion to the surface on which they are growing.
Some examples of positive signals:
- growth factors for neurons
- Interleukin-2 (IL-2), an essential factor for the mitosis of lymphocytes
Receipt of negative signals
- increased levels of oxidants within the cell
- damage to DNA by these oxidants or other agents like
- molecules that bind to specific receptors on the cell surface and signal the cell to begin the apoptosis program.
These death activators include:
- Tumor necrosis factor (TNF) that binds to the TNF receptor
- Lymphotoxin that also binds to the TNF receptor
- Fas ligand (FasL), a molecule that binds to a cell-surface receptor named Fas (also called CD95)
The Mechanisms of Apoptosis
There are 2 different mechanisms by which a cell commits suicide by apoptosis.
- one generated by signals arising within the cell
- the other triggered by death activators binding to receptors at the cell surface.
- TNF
- Lymphotoxin
- Fas ligand (FasL)
Apoptosis triggered by internal signals
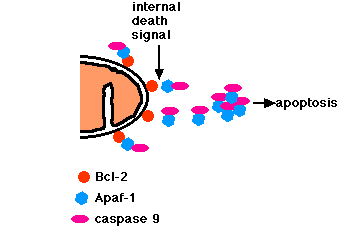
- In a healthy cell, the outer membranes of
- mitochondria
- the endoplasmic reticulum (ER)
- the nuclear envelope
- express the protein Bcl-2 on their surface.
- Bcl-2 binds a molecule of the protein
- Apaf-1, which is itself bound to a molecule of
- caspase 9
- Caspase 9 is one of a family of over a dozen caspases. They are all proteases. They get their name because they cleave proteins - mostly each other - at aspartic acid (Asp) residues).
- The trimer of Bcl-2/Apaf-1/caspase 9 has been tentatively named the apoptosome.
- Internal damage in the cell causes Bcl-2 to release the heterodimer of Apaf-1 and caspase 9.
- These aggregate in the cytosol.
- Caspase 9 cleaves and, in so doing, activates other caspases.
- The sequential activation of one caspase by another creates an expanding cascade of proteolytic activity (rather like that in blood clotting and complement activation) which leads to
- digestion of structural proteins in the cytoplasm
- degradation of chromosomal DNA and
- death of the cell
Apoptosis triggered by external signals
- Fas and the TNF receptor are integral membrane proteins with their receptor domains exposed at the surface of the cell
- binding of the complementary death activator (FasL and TNF respectively) transmits a signal to the cytoplasm that leads to
- activation of caspase 8
- caspase 8 (like caspase 9) initiates a cascade of caspase activation leading to
- death of the cell.
Apoptosis and Cancer
Some cancer-causing viruses use tricks to prevent apoptosis of the cells they have transformed.
- Two human papilloma viruses (HPV) have been implicated in causing cervical cancer. One of them produces a protein (E6) that binds and inactivates the apoptosis promoter p53.
- Epstein-Barr Virus (EBV), the cause of mononucleosis and a cause of Burkitt's lymphoma
- produces a protein similar to Bcl-2
- produces another protein that causes the cell to increase its own production of Bcl-2. Both these actions make the cell more resistant to apoptosis (thus enabling the cancer cell to continue to proliferate).
Even cancer cells produced without the participation of viruses may have tricks to avoid apoptosis.
- Some cancer cells produce high levels of Bcl-2, thus blocking apoptotic signals they may receive.
- melanoma is a notorious example
- Some B-cell leukemias and lymphomas express high levels of Bcl-2 because of a translocation of the BCL-2 gene into an enhancer region for antibody production. [Discussion].
- Some cancer cells express high levels of FasL, thus killing any cytotoxic T cells (CTL) that try to kill them. (CTL also express Fas; and this mechanism is the reverse of the one shown below).
- Other cancer cells, especially lung and colon cancer cells, secrete elevated levels of a soluble "decoy" molecule that binds to FasL, plugging it up so it cannot bind Fas. Thus, cytotoxic T cells (CTL) cannot kill the cancer cells by the mechanism shown below.
Apoptosis and AIDS
The hallmark of AIDS (acquired immunodeficiency syndrome) is the decline in the number of the patient's CD4+ T cells (normally about 1000 per microliter (µl) of blood). CD4+ T cells are responsible, directly or indirectly (as helper cells), for all immune responses. When their number declines below about 200 per µl, the patient is no longer able to mount effective immune responses and begins to suffer a series of dangerous infections.
What causes the disappearance of CD4+ T cells?
HIV (human immunodeficiency virus) invades CD4+ T cells by binding to their surface CD4 molecules. This binding is mediated by gp120, a surface glycoprotein (encoded by the virus's env gene). One might assume that it is infection by HIV that causes these cells to die. However, that appears not to the main culprit. Fewer than 1 in 100,000 CD4+ T cells in the blood of AIDS patients are actually infected with the virus.
So what kills so many uninfected CD4+ cells?
The answer is clear: apoptosis.
The mechanism is not clear. There are several possibilities.
One of them:
- gp120 is a peripheral membrane protein and much of it leaves the membrane and enters the blood and lymph in soluble form.
- soluble gp120 binds to the CD4 molecules on uninfected T cells
- (perhaps aided by the anti-gp120 antibodies that are produced following HIV infection and that are the basis of the most common blood test for HIV infection)
- this binding cross-links the CD4 molecules on the T cell leading to
- increased expression of Fas.
- If these cells encounter a cell expressing FasL (macrophages and cytotoxic T cells [right] can), or even begin expressing FasL themselves, they
- commit suicide by apoptosis.
For many years it has been known that certain parts of the body
- the anterior chamber of the eye
- the testes
are "immunologically privileged sites". Antigens within these sites fail to elicit an immune response.
It turns out that cells in these sites differ from the other cells of the body in that they express high levels of FasL at all times.
Thus antigen-reactive T cells, which express Fas, would be killed when they enter these sites.
This finding raises the possibility of a new way of preventing graft rejection.
If at least some of the cells on a transplanted kidney, liver, heart, etc. could be made to express high levels of FasL, that might protect the graft from attack by the T cells of the host's cell-mediated immune system. If so, then the present need for treatment with immunosuppressive drugs for the rest of the transplant recipient's life would be reduced or eliminated.
So far, the results in animal experiments have been mixed. Allografts engineered to express FasL have shown increased survival for kidneys but not for hearts or islets of Langerhans.
19 May 1999
{kind=link}