The ability of the immune system to respond to an antigen exists before it ever encounters that antigen.
The immune system relies on the prior formation of an incredibly diverse population of:| Link to discussion of how the diversity of BCRs and TCRs is created. |
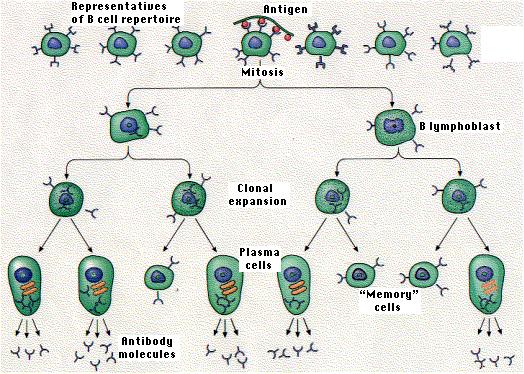
Each receptor is created even though the epitope it recognizes may never have been present in the body. If an antigen with that epitope should enter the body, those few lymphocytes able to bind to it will do so. If they also receive a second co-stimulatory signal, they may leave G0 of the cell cycle and begin repeated rounds of mitosis (now called lymphoblasts). In this way, clones of antigen-specific lymphocytes (B and T) develop providing the basis of the immune response.
The drawing illustrates the activation of the one B cell in a pool of B cells whose BCR is specific for an epitope (small dark spheres) on the antigen.This phenomenon is called clonal selection because it is antigen that selects particular lymphocytes for clonal expansion.
Clonal selection leads to the eventual production of: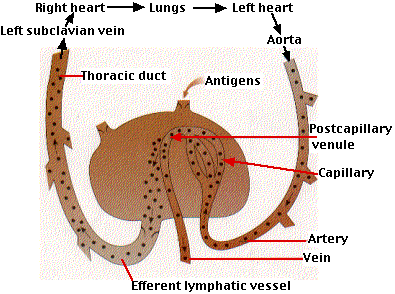
What is the probability that those few lymphocytes able to bind to a particular epitope will actually encounter the antigen carrying that epitope?
Surprisingly, it is quite high because both B cells and T cells migrate in and out of lymph nodes. Lymph nodes serve as lymph filters, trapping foreign matter that gains access to the body. (It has been shown that as much as 99% of the bacteria entering a node are removed by it.)
Lymph nodes, like any other tissue, receive a blood supply. When lymphocytes find themselves in a venule draining the capillaries of a lymph node (called a postcapillary venule), they migrate through the wall of the venule and take up temporary residence in the node. The unceasing traffic of B and T cells in and out of lymph nodes increases the probability that when an antigen enters the body, the appropriate B and T cells will encounter it.
The lymph draining from a node (into the efferent lymphatic vessel) contains a small number of B cells and a large number of T cells. These return to the blood at the subclavian veins. From there they are distributed throughout the body.
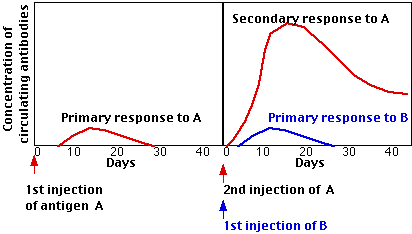
After recovering from an infection, the concentration of antibodies against the infectious agent gradually declines over the ensuing weeks, months, or even years. A time may come when antibodies against that agent can no longer be detected. Nevertheless, the individual often is still protected against a second case of the disease; that is, the person is still immune. In fact, a second exposure to the agent usually calls forth a more rapid and larger response to the antigen. This is called the secondary response.
The graphs demonstrate the main features of the secondary antibody response. A second injection of antigen A produces a secondary response. But the response is specific, not simply the result of a general enhancement of the immune system, because the simultaneous injection of a new antigen (B) produces only a primary response to it.
The secondary response reflects a larger number of antigen-specific cells - called memory cells - than existed before the primary response. During the initial expansion of clones, some of the progeny cells neither went on dividing nor developed into plasma cells. Instead, they reverted to small lymphocytes bearing the same BCR on their surface that their ancestors had. [look back at figure] This lays the foundation for a more rapid and massive response the next time the antigen enters the body.
To provide long-lasting immunity, memory cells must have long life spans if we assume that they never divide again unless they encounter the appropriate antigen. Some of the lymphocytes that circulate from lymph nodes to blood and back again do appear to be long-lived. In fact, there is evidence in humans that some small lymphocytes survive at least 20 years; perhaps for life.
| Links to discussions of |
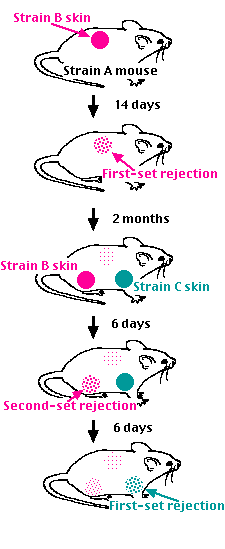
Graft rejection is a form of cell-mediated immunity. If a piece of skin from a mouse of one strain (B) is grafted onto the flank of a mouse of a second strain (A), the graft does well at first. Blood vessels from the host grow into it, and it functions normally. After some 10-14 days, however, the blood supply to the graft breaks down and the graft degenerates. Finally it is sloughed off like an old scab. This is called a first-set rejection.
That the graft rejection is an immune response is demonstrated by now grafting the mouse with two pieces of skin:Immunological memory provides the basis for the use of vaccines. These are preparations of infectious or toxic agents that have been altered so as not to cause disease. The alteration is not, however, so drastic as to destroy all the epitopes. The antibodies (and/or T cells) produced in response to the vaccine will also protect against the unaltered disease-producing agent.
| Link to a discussion of vaccines. |
| Welcome&Next Search |